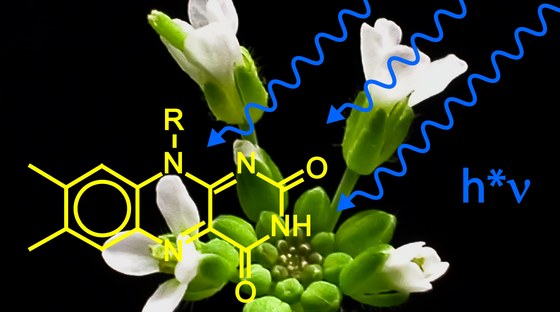
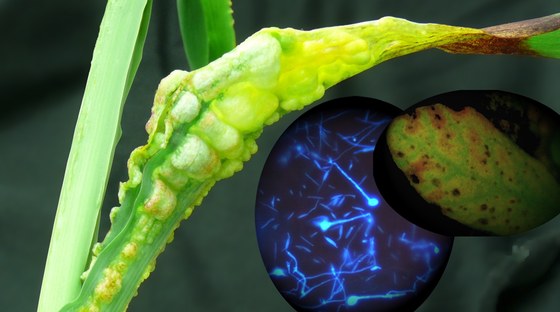
Willkommen im Fachgebiet Molekulare Pflanzenphysiologie
Molekulare Pflanzenphysiologie & Photobiologie (EN)
Molekulare Pflanzenphysiologie (EN)
Molekulare Pflanzenwissenschaften (EN)