Main Content
Bachelor's and Master's Theses
Table of Contents:
- Sensory motor component of reaction times
- Movement Primitives While Dancing
- Can people learn to control an additional, virtual arm?
- How immersive is consumer VR compared to high-end VR?
- Test of visual similarity models
Sensory motor component of reaction times
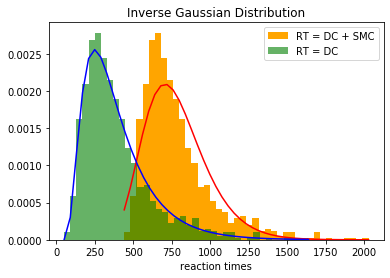
Goal: we want to build a comprehensive model of attentional guidance from behavioral data. To complete a part of the model, we are interested in disentangling the decision component and sensory motor component (SMC) of reaction times (see an example in Figure 1a). The missing part is that we do not confidently know how SMC is distributed. This is going to be investigated in this bachelor/master thesis.
Background: it is believed that total observed reaction time is a sum of different time components. As Luce mentioned [1] at least five parts can be included in a total reaction time: physical input changing into neural spike, spike transmission to the brain, signal processing and order making for a relevant muscle group (lets call this part decision time), signal transmission from the decision center to the muscles and eventually muscles reaction. As it is hard to observe all these components separately, we stack them all - except decision time - and call them here sensory motor component of reaction time.
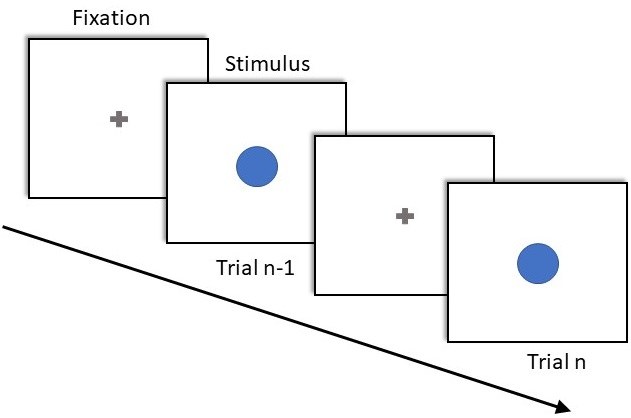
Your job: the main question of this project is to find out the distribution of sensory-motor component of reaction times by running an experiment where participants are asked to press the only available button as soon as they recognize the stimulus on the screen. This type of experiments are called simple response time (SRT) test in literature (see an example in FIgure 1b). Then we need to use machine learning methods to determine the best distribution by model selection. In some similar experiments the mean of simple reaction time is reported about: 208-224 ms [2], 194-256 ms [3] , 223-256 ms [4] and 213-231 ms [5] but the reaction time distribution is not described in these papers. The importance of knowing RT distributions is that this knowledge improves statistical analysis and, modeling cognitive and psychological processes. In some older research, such as [6], is reported that reaction time is a convolution of two Gaussian and exponential components, one represents the decision time and another represents motor component although some other researchers believed that it can not be convincingly concluded from the experiment (to follow the discussion see [1] page 102). Therefore, we will try to answer this question with a combination of psycho-physics and machine learning.
|
| (a) |
|
| (b) |
Figure 1: a) Reaction time distribution for a participant in a multi-forced-choice task. It is tried to find decision component (DC) of reaction times (RT) by fitting inverse Gaussian distribution. (b) Simple reaction time test
Literatur:
[1] R. D. Luce, Response Times: Their Role in Inferring Elementary Mental Organization. New York: Oxford University Press, 2008.
[2] R. Amini Vishteh, A. Mirzajani, E. Jafarzadehpour, and S. Darvishpour, "Evaluation of simple visual reaction time of dfferent colored light stimuli in visually normal students," Clinical Optometry, vol. 11, pp. 167-171, 2019.
[3] A. Batra, D. Jeph, and S. Vyas, "Color difference on simple visual reaction time in young volunteers," International Journal of Clinical and Experimental Physiology, vol. 1, no. 4, p. 311, 2014.
[4] A. Jain, R. Bansal, A. Kumar, and K. Singh, "A comparative study of visual and auditory reaction times on the basis of gender and physical activity levels of medical first year students," International Journal of Applied and Basic Medical Research, vol. 5, no. 2, p. 124, 2015.
[5] D. L. Woods, J. M. Wyma, E. W. Yund, T. J. Herron, and B. Reed, "Factors in uencing the latency of simple reaction time," Frontiers in Human Neuroscience, vol. 9, no. MAR, pp. 1-12, 2015.
[6] R. H. Hohle, "Inferred components of reaction times as functions of foreperiod duration," Journal of Experimental Psychology, vol. 69, no. 4, pp. 382-386, 1965.
Kontakt: Neda Meibodi / Prof. Dr. Dominik Endres
Movement Primitives While Dancing
Our bodies have a large number of joints that need to be controlled by our nervous systems in real time. A motor control theory postulates the existence of movement primitives to simplify this control. A movement primitive is a stored, stereotypical motion component that can be combined as needed in combination with other primitives to form new movements.
Whether these primitives exist, that is to say, they are implemented in the nervous system, is an unexplained question to this day. In this Bachelor / Master thesis it should be investigated if movement primitives are also suitable for modeling dance behavior. For this, we will record the dance movements of the subjects in the virtual reality & motion capture test room and model them using movement primitives. The movements thus modeled should then be validated in a perception experiment.
Literature:
D. Endres, E. Chiovetto, and M.A. Giese (2013). Model selection for the extraction of movement primitives. Frontiers in Computational Neuroscience, 7:185, 2013. DOI:10.3389/fncom.2013.00185
D. Endres, E. Chiovetto, and M.A. Giese (2015). Bayesian approaches for learning of primitive-based compact representations of complex human activities. In J.P. Laumond and A. Naoko, editors, Dance notation and robot motion, Springer Tracts in Advanced Robotics 111. Springer. DOI: 10.1007/978-3-319-25739-6_6
Contact: Prof. Dr. Dominik Endres
Can people learn to control an additional, virtual arm?
Our bodies have a large number of joints that need to be controlled by our nervous systems in real time. A theory for motor control postulates that this is done by optimal control, i. E. for each task goal, an optimal movement is calculated to achieve that goal. Two essential predictions of this theory are a) higher variance in degrees of freedom (joint angles) which are irrelevant to the current task than in task-relevant degrees of freedom, and b) the existence of synergies between degrees of freedom.
This project aims to investigate whether the task-irrelevant degrees of freedom with higher variance can be used to control a third, virtual arm, or whether this variance is not controllable. For this purpose, an experiment in the virtual reality test room will be performed, in which subjects receive a third, virtual arm.
First, the degrees of freedom of high variance are determined by motion recording, then these degrees of freedom are coupled to the third arm as a control scheme. The participants then have to learn a 3-arm manipulation task. We want to determine the learnability of the task, as well as quantify the difficulty of the learning task and examine the variance in the 'learned' degrees of freedom.
We also want to understand if the participants are learning new movement primitives/synergies for this task.
Literature:
Won, A. S., Bailenson, J., Lee, J., & Lanier, J. (2015). Homuncular flexibility in virtual reality. Journal of Computer-Mediated Communication, 20(3), 241-259. DOI: 10.1111/jcc4.12107
Todorov, E. (2004). Optimality principles in sensorimotor control. Nature neuroscience, 7(9), 907. DOI: 10.1038/nn1309
Endres, D. M., Chiovetto, E., & Giese, M. (2013). Model selection for the extraction of movement primitives. Frontiers in computational neuroscience, 7, 185. DOI: 10.3389/fncom.2013.00185
Contact: Prof. Dr. Dominik Endres
How immersive is consumer VR compared to high-end VR?
In the department's virtual reality testing room, subjects immerse themselves in virtual realities by wearing a head-mounted display (HMD) to create a three-dimensional, immersive virtual-reality experience. But how credible are these immersion experiences really? What is the difference between a (cheap) consumer-grade HMD and a much more expensive, professional HMD? We want to investigate this with the help of a rubber hand illusion: in the virtual realities, the test subjects are provided with a virtual body whose credibility is tested by means of questionnaires and reflex reactions. What interests us here is whether there is a measurable credibility difference between the Oculus Rift DK2 HMD, the HTC-Vive HMD and the NVIS ST50 HMD.
Literature:
Taubert, N., Löffler, M., Ludolph, N., Christensen, A., Endres, D., & Giese, M. A. (2013, August). A virtual reality setup for controllable, stylized real-time interactions between humans and avatars with sparse gaussian process dynamical models. In Proceedings of the ACM Symposium on Applied Perception (pp. 41-44). ACM. DOI: 10.1145/2492494.2492515
Slater, M., Spanlang, B., Sanchez-Vives, M. V., & Blanke, O. (2010). First person experience of body transfer in virtual reality. PloS one, 5(5), e10564. DOI:10.1371/journal.pone.0010564
Contact: Prof. Dr. Dominik Endres
Test of visual similarity models
(Visual) similarity is often used in cognitive neuroscience to compare mental and neuronal representations. For valid comparisons, a formalization of the concept of similarity is required, which can model incomparability and non-transitivity, since these properties are found in human data. With popular standard methods, such as multi-dimensional scaling and embedding in Euclidean spaces, these properties can not be derived, but A. Tversky's contrast model of similarity they can.
We want to investigate whether the axioms of this model in the expanded form according to Geist et. al (1996) depict the behavior of study participants. In particular, the question should be answered as to whether features of objects can be used to predict their pair suborder (see also one of our past empirical practica, in which we discussed this question but did not answer it).
Literature:
A. Tversky (1977) Features of similarity. Psychological Review, Vol 84(4), Jul 1977, 327-352. DOI: 10.1037/0033-295X.84.4.327
Geist, S., Lengnink, K., & Wille, R. (1996). An order-theoretic foundation for similarity measures (pp. 225-237). Springer Berlin Heidelberg. ISBN 3-8265-1393-2
Endres, D., Adam, R., Giese, M. A., & Noppeney, U. (2012, May). Understanding the semantic structure of human fMRI brain recordings with formal concept analysis. In International Conference on Formal Concept Analysis (pp. 96-111). Springer, Berlin, Heidelberg. DOI: 10.1007/978-3-642-29892-9_13
Contact: Prof. Dr. Dominik Endres